
Classificação dos Seres Vivos
A sistemática é a ciência dedicada a inventariar e descrever a biodiversidade e compreender as relações filogenéticas entre os organismos. Inclui a taxonomia (ciência da descoberta, descrição e classificação das espécies e grupo de espécies, com suas normas e princípios) e também a filogenia (relações evolutivas entre os organismos). Em geral, diz-se que compreende a classificação dos diversos organismos vivos. Em biologia, os sistematas são os cientistas que classificam as espécies em outros táxons a fim de definir o modo como eles se relacionam evolutivamente.
O objetivo da classificação dos seres vivos, chamada taxonomia, foi inicialmente o de organizar as plantas e animais conhecidos em categorias que pudessem ser referidas. Posteriormente a classificação passou a respeitar as relações evolutivas entre organismos, organização mais natural do que a baseada apenas em características externas. Para isso se utilizam também características ecológicas, fisiológicas, e todas as outras que estiverem disponíveis para os táxons em questão. É a esse conjunto de investigações a respeito dos táxons que se dá o nome de Sistemática. Nos últimos anos têm sido tentadas classificações baseadas na semelhança entre genomas, com grandes avanços em algumas áreas, especialmente quando se juntam a essas informações aquelas oriundas dos outros campos da Biologia.
A classificação dos seres vivos é parte da sistemática, ciência que estuda as relações entre organismos, e que inclui a coleta, preservação e estudo de espécimes, e a análise dos dados vindos de várias áreas de pesquisa biológica.
O primeiro sistema de classificação foi o de Aristóteles no século IV a.C., que ordenou os animais pelo tipo de reprodução e por terem ou não sangue vermelho. O seu discípulo Teofrasto classificou as plantas por seu uso e forma de cultivo.
Nos séculos XVII e XVIII os botânicos e zoólogos começaram a delinear o atual sistema de categorias, ainda baseados em características anatômicas superficiais. No entanto, como a ancestralidade comum pode ser a causa de tais semelhanças, este sistema demonstrou aproximar-se da natureza, e continua sendo a base da classificação atual. Lineu fez o primeiro trabalho extenso de categorização, em 1758, criando a hierarquia atual.
A partir de Darwin a evolução passou a ser considerada como paradigma central da Biologia, e com isso evidências da paleontologia sobre formas ancestrais, e da embriologia sobre semelhanças nos primeiros estágios de vida. No século XX, a genética e a fisiologia tornaram-se importantes na classificação, como o uso recente da genética molecular na comparação de códigos genéticos. Programas de computador específicos são usados na análise matemática dos dados.
Em fevereiro de 2005 Edward Osborne Wilson, professor aposentado da Universidade de Harvard, onde cunhou o termo biodiversidade e participou da fundação da sociobiologia, ao defender um "projeto genoma" da biodiversidade da Terra, propôs a criação de uma base de dados digital com fotos detalhadas de todas a espécies vivas e a finalização do projeto Árvore da vida. Em contraposição a uma sistemática baseada na biologia celular e molecular, Wilson vê a necessidade da sistemática descritiva para preservar a biodiversidade.
Do ponto de vista econômico, defendem Wilson, Peter Raven e Dan Brooks, a sistemática pode trazer conhecimentos úteis na biotecnologia, e na contenção de doenças emergentes. Mais da metade das espécies do planeta é parasita, e a maioria delas ainda é desconhecida.
De acordo com a classificação vigente as espécies descritas são agrupadas em gêneros. Os gêneros são reunidos, se tiverem algumas características em comum, formando uma família. Famílias, por sua vez, são agrupadas em uma ordem. Ordens são reunidas em uma classe. Classes de seres vivos são reunidas em filos. E os filos são, finalmente, componentes de alguns dos cinco reinos (Monera, Protista, Fungi, Plantae e Animalia).

Nomenclatura Científica
Nomenclatura é a atribuição de nomes (nome científico) a organismos e às categorias nas quais são classificados.
O nome científico é aceito em todas as línguas, e cada nome aplica-se apenas a uma espécie.
Há duas organizações internacionais que determinam as regras de nomenclatura, uma para zoologia e outra para botânica. Segundo as regras, o primeiro nome publicado (a partir do trabalho de Lineu) é o correto, a menos que a espécie seja reclassificada, por exemplo, em outro gênero. A reclassificação tem ocorrido com certa frequência desde o século XX. O Código Internacional de Nomenclatura Zoológica preconiza que neste caso mantém-se a referência a quem primeiro descreveu a espécie, com o ano da decisão, entre parênteses, e não inclui o nome de quem reclassificou. Esta norma internacional decorre, entre outras coisas, do fato de ser ainda nova a abordagem genética da taxonomia, sujeita a revisão devido a novas pesquisas científicas, ou simplesmente a definição de novos parâmetros para a delimitação de um táxon, que podem ser morfológicos, ecológicos, comportamentais etc.
O sistema atual identifica cada espécie por dois nomes em latim: o primeiro, em maiúscula, é o gênero, o segundo, em minúscula, é o epíteto específico. Os dois nomes juntos formam o nome da espécie. Os nomes científicos podem vir do nome do cientista que descreveu a espécie, de um nome popular desta, de uma característica que apresente, do lugar onde ocorre, e outros. Por convenção internacional, o nome do gênero e da espécie é impresso em itálico, grifado ou em negrito, o dos outros táxons não. Subespécies têm um nome composto por três palavras.
Ex.: Canis familiares, Canis lupus, Felis catus.
Nomenclatura popular
A nomeação dos seres vivos que compõe a biodiversidade constitui uma etapa do trabalho de classificação. Muitos seres são "batizados" pela população com nomes denominados populares ou vulgares, pela comunidade científica.
Esses nomes podem designar um conjunto muito amplo de organismos, incluindo, algumas vezes, até grupos não aparentados.
O mesmo nome popular pode ser atribuído a diferentes espécies, como neste exemplo:
 Ananas comosus |  Ananas ananassoides |
Estas duas espécies do gênero ananas são chamadas pelo mesmo nome popular Abacaxi.
Outro exemplo é o crustáceo de praia Emerita brasiliensis, que no Rio de Janeiro é denominado tatuí, e nos estados de São Paulo e Paraná é chamado de tatuíra.

Em contra partida, animais de uma mesma espécie podem receber vários nomes, como ocorre com a onça-pintada, cujo nome científico é Panthera onca.

Outros nomes populares:
canguçu, onça-canguçu, jaguar-canguçu
Um outro exemplo é a planta Manihot esculenta, cuja raiz é muito apreciada como alimento. Dependendo da região do Brasil, ela é conhecida por vários nomes: aimpim, macaxeira ou mandioca.
Considerando os exemplo apresentados, podemos perceber que a nomenclatura popular varia bastante, mesmo num país como o Brasil, em que a população fala um mesmo idioma, excetuando-se os idiomas indígenas. Imagine se considerarmos o mundo todo, com tantos, com tantos idiomas e dialetos diferentes, a grande quantidade de nomes de um mesmo ser vivo pode receber. Desse modo podemos entender a necessidade de existir uma nomenclatura padrão, adotada internacionalmente, para facilitar a comunicação de diversos profissionais, como os médicos, os zoólogos, os botânicos e todos aqueles que estudam os seres vivos.
A Filogênese dos Seres Vivos
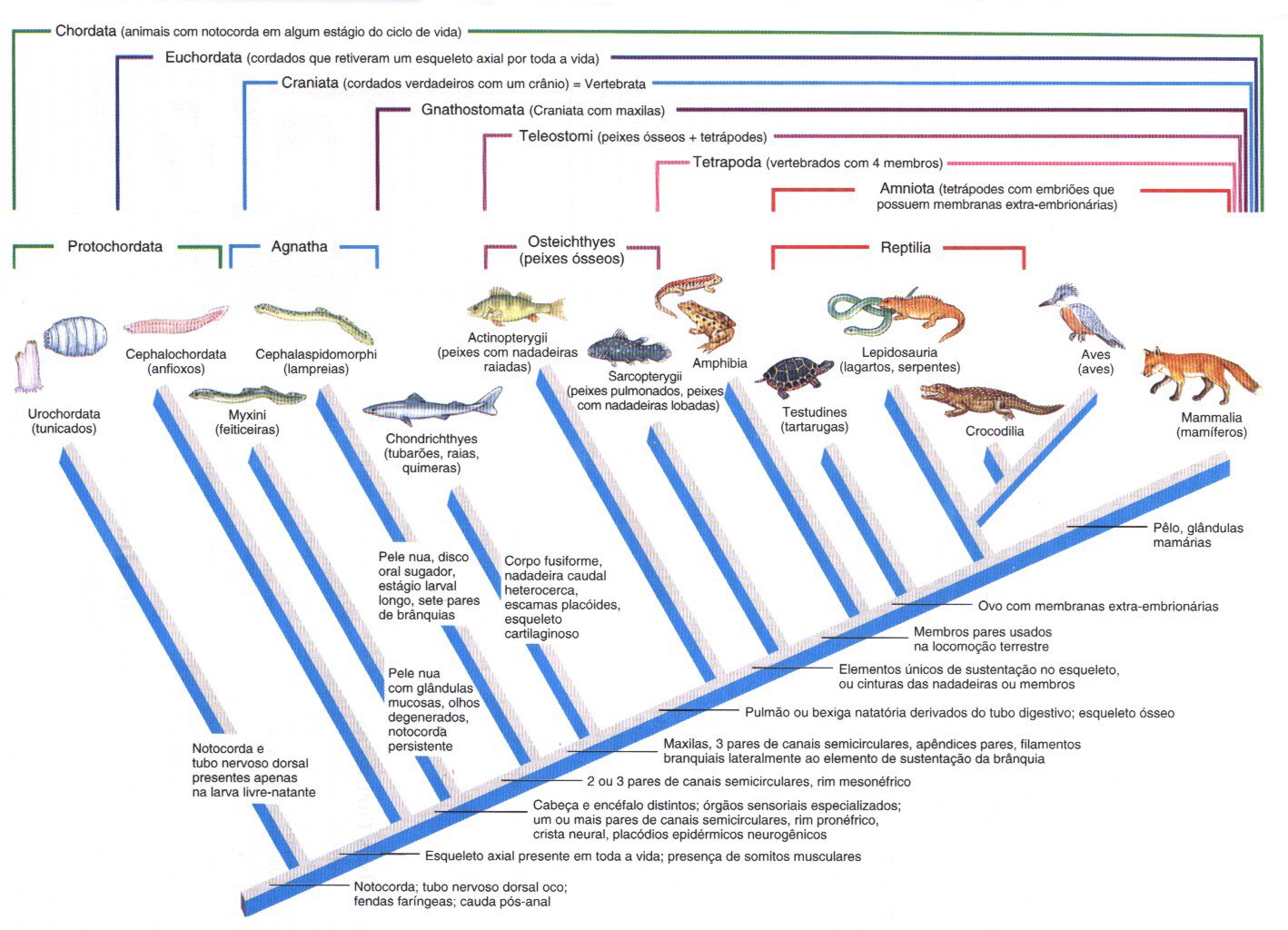
Qual foi o ancestral dos répteis (lagartos, cobras) que vivem na Terra atual? Essas e outras perguntas relativas à origem dos grandes grupo de seres vivos eram difíceis de serem respondidas até surgir, em 1959, a Teoria da evolução Biológica por Seleção Natural, proposta por Charles Darwin e Alfred Russel Wallace. Com a compreensão de "como" a evolução biológica ocorre, os biólogos passaram a sugerir hipóteses para explicar a possível relação de parentesco entre os diversos grupos de seres vivos.
Diagramas em forma de árvore - elaborados com dados de anatomia e embriologia comparadas, além de informações derivadas do estudo de fósseis - mostraram a hipotética origem de grupos a partir de supostos ancestrais. Essas supostas "árvores genealógicas" ou "filogenéticas" (do grego, phylon = raça, tribo + génesis = fonte, origem, início) simbolizavam a história evolutiva dos grupos que eram comparados, além de sugerir uma provável época de origem para cada um deles. Como exemplo veja a figura abaixo.

O esquema representa uma provável "história evolutiva" dos vertebrados. Note que estão representados os grupos atuais - no topo do esquema- bem como os prováveis ancestrais. Perceba que o grupo das lampreias (considerados "peixes" sem mandíbula) é bem antigo (mais de 500 milhões de anos). Já cerca de 150 milhões de anos, provavelmente a partir de um grupo de dinossauros ancestrais. Note, ainda, que o parentesco existe entre aves e répteis é maior do que existe entre mamífero e répteis, e que os três grupos foram originados de um ancestral comum.
Atualmente com um maior número de informações sobre os grupos taxonômicos passaram-se a utilizar computadores para se gerar as arvores filogenéticas e os cladogramas para estabelecer as inúmeras relações entre os seres vivos.
Estabelecendo Filogenias com os Cladogramas
Ao dispor de um grande número de características comparativas, mais confiáveis - anatômicas, embriológicas, funcionais, genéticas, comportamentais etc. - os biólogos interessados na classificação dos seres vivos puderam elaborar hipóteses mais consistentes a respeito da evolução dos grandes grupos. Influenciados pelo trabalho de Wili Hennig - um cientista alemão, especialista em insetos - passaram a apresentar as características em cladogramas. Neste tipo de diagrama, utiliza-se uma linha, cujo ponto de origem - a raiz- simboliza um provável grupo (ou espécie) ancestral. De cada nó surge um ramo, que conduz a um ou a vários grupos terminais. Com os cladogramas pode-se estabelecer uma comparação entre as características primitivas - que existiam em grupos ancestrais - e as derivadas - compartilhadas por grupos que os sucederam.
Vírus
Vírus são os únicos organismos acelulares da Terra atual.
Os vírus são seres muito simples e pequenos (medem menos de 0,2 µm), formados basicamente por uma cápsula protéica envolvendo o material genético, que, dependendo do tipo de vírus, pode ser o DNA, RNA ou os dois juntos (citomegalovirus). A palavra vírus vem do Latim virus que significa fluído venenoso ou toxina. Atualmente é utilizada para descrever os vírus biológicos, além de designar, metaforicamente, qualquer coisa que se reproduza de forma parasitária, como idéias. O termo vírus de computador nasceu por analogia. A palavra vírion ou víron é usada para se referir a uma única partícula viral que estiver fora da célula hospedeira.
Das 1.739.600 espécies de seres vivos conhecidos, os vírus representam 3.600 espécies.
 Ilustração do vírus HIV mostrando as proteínas do capsídeo responsáveis pela aderencia na célula hospedeira. | Vírus é uma partícula basicamente protéica que pode infectar organismos vivos. Vírus são parasitas obrigatórios do interior celular e isso significa que eles somente se reproduzem pela invasão e possessão do controle da maquinaria de auto-reprodução celular. O termo vírus geralmente refere-se às partículas que infectam eucariontes (organismos cujas células têm carioteca), enquanto o termo bacteriófago ou fago é utilizado para descrever aqueles que infectam procariontes (domínios bacteria e archaea). Tipicamente, estas partículas carregam uma pequena quantidade de ácido nucléico (seja DNA ou RNA, ou os dois) sempre envolto por uma cápsula protéica denominada capsídeo. As proteínas que compõe o capsídeo são específicas para cada tipo de vírus. O capsídeo mais o ácido nucléico que ele envolve são denominados nucleocapsídeo. Alguns vírus são formados apenas pelo núcleo capsídeo, outros no entanto, possuem um envoltório ou envelope externo ao nucleocapsídeo. Esses vírus são denominados vírus encapsulados ou envelopados. |
O envelope consiste principalmente em duas camadas de lipídios derivadas da membrana plasmática da célula hospedeira e em moléculas de proteínas virais, específicas para cada tipo de vírus, imersas nas camadas de lipídios.
São as moléculas de proteínas virais que determinam qual tipo de célula o vírus irá infectar. Geralmente, o grupo de células que um tipo de vírus infecta é bastante restrito. Existem vírus que infectam apenas bactérias, denominadas bacteriófagos, os que infectam apenas fungos, denominados micófagos; os que infectam as plantas e os que infectam os animais, denominados, respectivamente, vírus de plantas e vírus de animais.

Esquema do Vírus HIV
Os vírus não são constituídos por células, embora dependam delas para a sua multiplicação. Alguns vírus possuem enzimas. Por exemplo o HIV tem a enzima Transcriptase reversa que faz com que o processo de Transcrição reversa seja realizado (formação de DNA a partir do RNA viral). Esse processo de se formar DNA a partir de RNA viral é denominado retrotranscrição, o que deu o nome retrovírus aos vírus que realizam esse processo. Os outros vírus que possuem DNA fazem o processo de transcrição (passagem da linguagem de DNA para RNA) e só depois a tradução. Estes últimos vírus são designados de adenovírus.
Vírus são parasitas intracelulares obrigatórios: a falta de hialoplasma e ribossomos impede que eles tenham metabolismo próprio. Assim, para executar o seu ciclo de vida, o vírus precisa de um ambiente que tenha esses componentes. Esse ambiente precisa ser o interior de uma célula que, contendo ribossomos e outras substâncias, efetuará a síntese das proteínas dos vírus e, simultaneamente, permitirá que ocorra a multiplicação do material genético viral.
Em muitos casos os vírus modificam o metabolismo da célula que parasitam, podendo provocar a sua degeneração e morte. Para isso, é preciso que o vírus inicialmente entre na célula: muitas vezes ele adere à parede da célula e "injeta" o seu material genético ou então entra na célula por englobamento - por um processo que lembra a fagocitose, a célula "engole" o vírus e o introduz no seu interior.
Virus seres vivos ou não?
Vírus não têm qualquer atividade metabólica quando fora da célula hospedeira: eles não podem captar nutrientes, utilizar energia ou realizar qualquer atividade biossintética. Eles obviamente se reproduzem, mas diferentemente de células, que crescem, duplicam seu conteúdo para então dividir-se em duas células filhas, os vírus replicam-se através de uma estratégia completamente diferente: eles invadem células, o que causa a dissociação dos componentes da partícula viral; esses componentes então interagem com o aparato metabólico da célula hospedeira, subvertendo o metabolismo celular para a produção de mais vírus.
Há grande debate na comunidade científica sobre se os vírus devem ser considerados seres vivos ou não, e esse debate e primariamente um resultado de diferentes percepções sobre o que vem a ser vida, em outras palavras, a definição de vida. Aqueles que defendem a idéia que os vírus não são vivos argumentam que organismos vivos devem possuir características como a habilidade de importar nutrientes e energia do ambiente, devem ter metabolismo (um conjunto de reações químicas altamente inter-relacionadas através das quais os seres vivos constroem e mantêm seus corpos, crescem e performam inúmeras outras tarefas, como locomoção, reprodução, etc.); organismos vivos também fazem parte de uma linhagem continua, sendo necessariamente originados de seres semelhantes e, através da reprodução, gerar outros seres semelhantes (descendência ou prole), etc.
Os vírus preenchem alguns desses critérios: são parte de linhagens contínuas, reproduzem-se e evoluem em resposta ao ambiente, através de variabilidade e seleção, como qualquer ser vivo. Porém, não têm metabolismo próprio, por isso deveriam ser considerados "partículas infecciosas", ao invés de seres vivos propriamente ditos. Muitos, porém, não concordam com essa perspectiva, e argumentam que uma vez que os vírus são capazes de reproduzir-se, são organismos vivos; eles dependem do maquinário metabólico da célula hospedeira, mas até ai todos os seres vivos dependem de interações com outros seres vivos. Outros ainda levam em consideração a presença massiva de vírus em todos os reinos do mundo natural, sua origem-aparentemente tão antiga como a própria vida - sua importância na história natural de todos os outros organismos, etc. Conforme já mencionado, diferentes conceitos a respeito do que vem a ser vida formam o cerne dessa discussão. Definir vida tem sido sempre um grande problema, e já que qualquer definição provavelmente será evasiva ou arbitraria, dificultando assim uma definição exata a respeito dos vírus.
Doenças humanas virais
No homem, inúmeras doenças são causadas por esses seres acelulares. Praticamente todos os tecidos e órgãos humanos são afetados por alguma infecção viral. Abaixo você encontra as viroses mais frequentes na nossa espécie. Valorize principalmente os mecanismos de transmissão e de prevenção. Note que a febre amarela e dengue são duas viroses que envolvem a transmissão por insetos (mosquito da espécie Aedes aegypti). Para a primeira, existe vacina. Duas viroses relatadas abaixo, AIDS e condiloma acuminado, são doenças sexualmente trasmissíveis (DSTs). A tabela também relaciona viroses comuns na infância, rubélola, caxumba, sarampo, poliomelite - para as quais exiestem vacinas.
Algumas das principais viroses que acometem os seres humanos:
- Resfriado Comum;
- Caxumba;
- Raiva;
- Rubéola;
- Sarampo;
- Hepatites;
- Dengue;
- Poliomielite;
- Febre amarela;
- Varicela ou Catapora;
- Varíola;
- Meningite viral;
- Mononucleose Infecciosa;
- Herpes
- Condiloma
- Hantavirose
- AIDS.
Prevenção e tratamento de doenças virais
Devido ao uso da maquinaria das células do hospedeiro, os vírus tornam-se difíceis de matar. As mais eficientes soluções médicas para as doenças virais são, até agora, as vacinas para prevenir as infecções, e drogas que tratam os sintomas das infecções virais. Os pacientes frequentemente pedem antibióticos, que são inúteis contra os vírus, e seu abuso contra infecções virais é uma das causas de resistência antibiótica em bactérias. Diz-se, às vezes, que a ação prudente é começar com um tratamento de antibióticos enquanto espera-se pelos resultados dos exames para determinar se os sintomas dos pacientes são causados por uma infecção por vírus ou bactérias.
Bacteriófagos
Os bacteriófagos podem ser vírus de DNA ou de RNA que infectam somente organismos procariotos. São formados apenas pelo nucleocapsídeo, não existindo formas envelopadas. Os mais estudados são os que infectam a bactéria intestinal Escherichia coli, conhecida como fagos T. Estes são constituídos por uma cápsula protéica bastante complexa, que apresenta uma região denominada cabeça, com formato poligonal, envolvendo uma molécula de DNA, e uma região denominada cauda, com formato cilíndrico, contendo, em sua extremidade livre, fibras protéicas.
A reprodução ou replicação dos bacteriófagos, assim como os demais vírus, ocorre somente no interior de uma célula hospedeira.
Existem basicamente dois tipos de ciclos reprodutivos: o ciclo lítico e o ciclo lisogênico. Esses dois ciclos iniciam com o fago T aderindo à superfície da célula bacteriana através das fibras protéicas da cauda. Esta contrai-se, impelindo a parte central, tubular, para dentro da célula, à semelhança, de uma microsseringa. O DNA do vírus é, então, injetado fora da célula a cápsula protéica vazia. A partir desse momento, começa a diferenciação entre ciclo lítico e ciclo lisogênico.
No ciclo lítico, o vírus invade a bactéria, onde as funções normais desta são interrompidas na presença de ácido nucléico do vírus (DNA ou RNA). Esse, ao mesmo tempo em que é replicado, comanda a síntese das proteínas que comporão o capsídeo. Os capsídeos organizam-se e envolvem as moléculas de ácido nucléico. São produzidos, então novos vírus. Ocorre a lise, ou seja, a célula infectada rompe-se e os novos bacteriófagos são liberados. Sintomas causados por um vírus que se reproduz através desta maneira, em um organismo multicelular aparecem imediatamente. Nesse ciclo, os vírus utilizam o equipamento bioquímico(Ribossomo)da célula para fabricar sua proteína (Capsídeo). No ciclo lisogênico, o vírus invade a bactéria ou a célula hospedeira, onde o DNA viral incorpora-se ao DNA da célula infectada. Isto é, o DNA viral torna-se parte do DNA da célula infectada. Uma vez infectada, a célula continua suas operações normais, como reprodução e ciclo celular. Durante o processo de divisão celular, o material genético da célula, juntamente com o material genético do vírus que foi incorporado, sofrem duplicação e em seguida são divididos equitativamente entre as células-filhas. Assim, uma vez infectada, uma célula começará a transmitir o vírus sempre que passar por mitose e todas as células estarão infectadas também. Sintomas causados por um vírus que se reproduz através desta maneira, em um organismo multicelular podem demorar a aparecer. Doenças causadas por vírus lisogênico tendem a ser incuráveis. Alguns exemplos incluem a AIDS e herpes. |  |
Sob determinadas condições, naturais e artificiais (tais como radiações ultravioleta, raios X ou certos agentes químicas), uma bactéria lisogênica pode transformar-se em não-lisogênica e iniciar o ciclo lítico.
Reino Monera
O reino monera é formado por bactérias, cianobactérias e arqueobactérias (também chamadas arqueas), todos seres muito simples, unicelulares e com célula procariótica (sem núcleo diferenciado). Esses seres microscópios são geralmente menores do que 8 micrômetros ( 1µm = 0,001 mm).
As bactérias (do grego bakteria: 'bastão') são encontrados em todos os ecossistemas da Terra e são de grande importância para a saúde, para o ambiente e a economia. As bactérias são encontradas em qualquer tipo de meio: mar, água doce, solo, ar e, inclusive, no interior de muitos seres vivos.
Exemplos da importância das bactérias:
- na decomposição de matéria orgânica morta. Esse processo é efetuado tanto aeróbia, quanto anaerobiamente;
- agentes que provocam doença no homem;
- em processos industriais, como por exemplo, os lactobacilos, utilizados na indústria de transformação do leite em coalhada;
- no ciclo do nitrogênio, em que atuam em diversas fases, fazendo com que o nitrogênio atmosférico possa ser utilizado pelas plantas;
- em Engenharia Genética e Biotecnologia para a síntese de várias substâncias, entre elas a insulina e o hormônio de crescimento.
Estrutura das Bactérias
Bactérias são microorganismos unicelulares, procariotos, podendo viver isoladamente ou construir agrupamentos coloniais de diversos formatos. A célula bacterianas contém os quatro componentes fundamentais a qualquer célula: membrana plasmática, hialoplasma, ribossomos e cromatina, no caso, uma molécula de DNA circular, que constitui o único cromossomo bacteriano.
A região ocupada pelo cromossomo bacteriano costuma ser denominada nucleóide. Externamente à membrana plasmática existe uma parede celular (membrana esquelética, de composição química específica de bactérias).
É comum existirem plasmídios - moléculas de DNA não ligada ao cromossomo bacteriano - espalhados pelo hialoplasma. Plasmídios costumam conter genes para resistência a antibióticos.

Algumas espécies de bactérias possuem, externamente à membrana esquelética, outro envoltório, mucilaginoso, chamado de cápsula. É o caso dos pneumococos (bactérias causadoras de pneumonia). Descobriu-se que a periculosidade dessas bactérias reside na cápsula em um experimento, ratos infectados com pneumococo sem cápsula tiveram a doença porém não morreram, enquanto pneumococos capsulados causaram pneumonia letal.
A parede da célula bacteriana, também conhecida como membrana esquelética, reveste externamente a membrana plasmática, e é constituída de uma substância química exclusiva das bactérias conhecida como mureína (ácido n-acetil murâmico).
A Diversidade Metabólica das Bactérias
Se há um grupo de seres que apresenta grande diversidade metabólica, certamente é o das bactérias. Existem espécies heterótrofas e espécies autótrofas. Dentre as primeiras, destacam-se as parasitas, as decompositoras de matéria orgânica e as que obtêm matéria orgânica de outros seres vivos, com os quais se associam sem prejudicá-los. Dentre as autótrofas, existem espécies que produzem matéria orgânica por fotossíntese e outras que produzem por quimiossíntese.
As bactérias Heterótrofas
As bactérias parasitas são as que, por meio de inúmeros mecanismos, agridem outros seres vivos para a obtenção de alimento orgânico e causam inúmeras doenças. As decompositoras (frequentemente denominadas sapróvoras, saprofíticas ou saprofágicas) obtêm o alimento orgânico recorrendo à decomposição da matéria orgânica morta e são importântes na reciclagem dos nutrientes minerais na biosfera.
As que são associadas as outros seres vivos são denominadas de simbiontes, e não agridem os parceiros. É o caso das bactérias encontradas no estômago dos ruminantes (bois, cabras), que se nutrem da celulose ingerida por esses animais, fornecendo, em troca, aminoácidos essenciais para o metabolismo protéico do mesmo.
Muitas bactérias heterótrofas são anaeróbias obrigatórias, como o bacilo do tétano. São bactérias que morrem na presença de oxigênio. Nesse caso a energia dos compostos orgânicos é obtida por meio de fermentação. As anaeróbicas facultativas, por outro lado, vivem tanto na presença como na ausência de oxigênio.
Outras espécies só sobrevivem em presença de oxigênio - são as aeróbias obrigatórias. Um curioso grupo de bactérias é o que realiza a respiração aeróbia. Nessa modalidade de metabolismo energético existem todas as etapas típicas da respiração celular. Muda apenas o aceptor final de elétrons na cadeia respiratória. No lugar do oxigênio, essas bactérias utilizam nitrato, nitrito ou sulfato, obtendo no final, praticamente o mesmo rendimento energético verificado na respiração celular aeróbia. É o que ocorre com as bactérias desnitrificantes que participam do ciclo do nitrogênio na natureza. Nelas o aceptor final de elétrons é o nitrato.
Bactérias Autótrofas
Fotossintetizantes
Nas bactérias que realizam fotossíntese, a captação da energia solar fica a cargo de uma clorofila conhecida como bacterioclorofila. A partir da utilização de substâncias simples do meio, ocorre a síntese do combustível biológico. De maneira geral, não há liberação de oxigênio. Como exemplo, podemos citar as bactérias sulforosas do gênero Chlorobium, que efetuam esse processo com a utilização de H2S e CO2, segundo a equação:
2H2S + CO2 + luz ------bacterioclorofila------------> (CH2) + 2S + H20
Note que é o gás sulfídrico, e não a água, que atua como fornecedor dos hidrogênios que servirão para a redução do gás carbônico. Não há a liberação de oxigênio. O enxofre permanece no interior das células bacterianas sendo, posteriormente eliminado para o meio em que vivem esses microorganismos, em geral fontes sulfurosas. Nesse processo, CH2O representa a matéria orgânica produzida.
Quimiossíntese
A quimiossíntese é uma reação que produz energia química, convertida da energia de ligação dos compostos inorgânicos oxidados. Sendo a energia química liberada, empregada na produção de compostos orgânicos e gás oxigênio (O2), a partir da reação entre o dióxido de carbono (CO2) e água molecular (H2O), conforme demonstrado abaixo:
- Primeira etapa
- Primeira etapa
Composto Inorgânico + O2 → Compostos Inorgânicos oxidados + Energia Química
- Segunda etapa
CO2 + H2O + Energia Química → Compostos Orgânicos + O2
Esse processo autotrófico de síntese de compostos orgânicos ocorre na ausência de energia solar. É um recurso normalmente utilizado por algumas espécies de bactérias e arqueobactérias (bactérias com características primitivas ainda vigentes), recebendo a denominação segundo os compostos inorgânicos reagentes, podendo ser: ferrobactérias e nitrobactérias ou nitrificantes (nitrossomonas e nitrobacter, gênero de bactérias quimiossíntetizantes).
As ferrobactérias oxidam substâncias à base de ferro para conseguirem energia química, já as nitrificantes, utilizam substâncias à base de nitrogênio.
Presentes no solo, as nitrossomonas e nitrobacter, são importantes organismos considerados biofixadores de nitrogênio, geralmente encontradas livremente no solo ou associadas às plantas, formando nódulos radiculares.
A biofixação se inicia com a assimilação no nitrogênio atmosférico (N2), transformando-o em amônia (NH3), reagente oxidado pela nitrossomona, resultando em nitrito (NO2-) e energia para a produção de substâncias orgânicas sustentáveis a esse gênero de bactérias.
O nitrito, liberado no solo e absorvido pela nitrobacter, também passa por oxidação, gerando energia química destinada à produção de substâncias orgânicas a esse gênero e nitrato (NO3-), aproveitado pelas plantas na elaboração dos aminoácidos.
Reação quimiossintética nas Nitrossomonas:
As ferrobactérias oxidam substâncias à base de ferro para conseguirem energia química, já as nitrificantes, utilizam substâncias à base de nitrogênio.
Presentes no solo, as nitrossomonas e nitrobacter, são importantes organismos considerados biofixadores de nitrogênio, geralmente encontradas livremente no solo ou associadas às plantas, formando nódulos radiculares.
A biofixação se inicia com a assimilação no nitrogênio atmosférico (N2), transformando-o em amônia (NH3), reagente oxidado pela nitrossomona, resultando em nitrito (NO2-) e energia para a produção de substâncias orgânicas sustentáveis a esse gênero de bactérias.
O nitrito, liberado no solo e absorvido pela nitrobacter, também passa por oxidação, gerando energia química destinada à produção de substâncias orgânicas a esse gênero e nitrato (NO3-), aproveitado pelas plantas na elaboração dos aminoácidos.
Reação quimiossintética nas Nitrossomonas:
NH3 (amônia) + O2 → NO2- (nitrito) + Energia
6 CO2 + 6 H2O + Energia → C6H12O6 (Glicose - Compostos Orgânicos) + 6 O2
6 CO2 + 6 H2O + Energia → C6H12O6 (Glicose - Compostos Orgânicos) + 6 O2
Reação quimiossintética nas Nitrobacter:
NO2- (nitrito) + O2 → NO3- (nitrato) + Energia
6 CO2 + 6 H2O + Energia → C6H12O6 + 6 O2
6 CO2 + 6 H2O + Energia → C6H12O6 + 6 O2
Assim, podemos perceber que o mecanismo de quimiossíntese, extremamente importante para a sobrevivência das bactérias nitrificantes, também é bastante relevante ao homem. Conforme já mencionado, o nitrito absorvido pelas plantas, convertidos em aminoácidos, servem como base de aminoácidos essenciais à nutrição do homem (um ser onívoro: carnívoro e herbívoro).
Dessa forma, fica evidente a interdependência existente entre os fatores bióticos (a diversidade dos organismos) e os fatores abióticos (aspectos físicos e químicos do meio ambiente).
Dessa forma, fica evidente a interdependência existente entre os fatores bióticos (a diversidade dos organismos) e os fatores abióticos (aspectos físicos e químicos do meio ambiente).
Reprodução das Bactérias
A reprodução mais comum nas bactérias é assexuada por bipartição ou cissiparidade. Ocorre a duplicação do DNA bacteriano e uma posterior divisão em duas células. As bactérias multiplicam-se por este processo muito rapidamente quando dispõem de condições favoráveis (duplica em 20 minutos). A separação dos cromossomos irmãos conta com a participação dos mesossomos, pregas internas da membrana plasmática nas quais existem também as enzimas participantes da maior parte da respiração celular. |  |
Repare que não existe a formação do fuso de divisão e nem de figuras clássicas e típicas da mitose. Logo, não é mitose.

Esporulação
Algumas espécies de bactérias originam, sob certas condições ambientais, estruturas resistentes denominadas esporos. A célula que origina o esporo se desidrata, forma uma parede grossa e sua atividade metabólica torna-se muito reduzida. Certos esporos são capazes de se manter em estado de dormência por dezenas de anos. Ao encontrar um ambiente adequado, o esporo se reidrata e origina uma bactéria ativa, que passa a se reproduzir por divisão binária.
Os esporos são muito resistentes ao calor e, em geral, não morrem quando expostos à água em ebulição. Por isso os laboratórios, que necessitam trabalhar em condições de absoluta assepsia, costumam usar um processo especial, denominado autoclavagem, para esterilizar líquidos e utensílios. O aparelho onde é feita a esterilização, a autoclave, utiliza vapor de água a temperaturas da ordem de 120ºC, sob uma pressão que é o dobro da atmosférica. Após 1 hora nessas condições, mesmo os esporos mais resistentes morrem.
A indústria de enlatados toma medidas rigorosas na esterilização dos alimentos para eliminar os esporos da bactéria Clostridium botulinum. Essa bactéria produz o botulismo, infecção frequentemente fatal.
Reprodução sexuada
Para as bactérias considera-se reprodução sexuada qualquer processo de transferência de fragmentos de DNA de uma célula para outra. Depois de transferido, o DNA da bactéria doadora se recombina com o da receptora, produzindo cromossomos com novas misturas de genes. Esses cromossomos recombinados serão transmitidos às células-filhas quando a bactéria se dividir.
A transferência de DNA de uma bactéria para outra pode ocorrer de três maneiras: por transformação, transdução e por conjugação.
Transformação
Na transformação, a bactéria absorve moléculas de DNA dispersas no meio e são incorporados à cromatina. Esse DNA pode ser proveniente, por exemplo, de bactérias mortas. Esse processo ocorre espontaneamente na natureza.
Os cientistas têm utilizado a transformação como uma técnica de Engenharia Genética, para introduzir genes de diferentes espécies em células bacterianas.

Transdução
Na transdução, moléculas de DNA são transferidas de uma bactéria a outra usando vírus como vetores (bactériófagos). Estes, ao se montar dentro das bactérias, podem eventualmente incluir pedaços de DNA da bactéria que lhes serviu de hospedeira. Ao infectar outra bactéria, o vírus que leva o DNA bacteriano o transfere junto com o seu. Se a bactéria sobreviver à infecção viral, pode passar a incluir os genes de outra bactéria em seu genoma.

Conjugação
Na conjugação bacteriana, pedaços de DNA passam diretamente de uma bactéria doadora, o "macho", para uma receptora, a "fêmea". Isso acontece através de microscópicos tubos protéicos, chamados pili, que as bactérias "macho" possuem em sua superfície.
O fragmento de DNA transferido se recombina com o cromossomo da bactéria "fêmea", produzindo novas misturas genéticas, que serão transmitidas às células-filhas na próxima divisão celular.

Conjugação bacterian mostrando o pili sexual.

As Cianobactérias
 | Extremamente parecidas com as bactérias, as cianobactérias são também procariontes. São todas autótrofas fotossintetizantes, mas suas células não possuem cloroplastos. A clorofila, do tipo a, fica dispersa pelo hialoplasma e em lamelas fotossintetizantes, que são ramificações da membrana plasmática. Além da clorofila, possuem outros pigmentos acessórios, como os carotenóides (pigmentos semelhantes ao caroteno da cenoura), ficoeritrina (um pigmento de cor vermelha, típico das cianobactérias encontradas no Mar vermelho) e a ficocianina (um pigmento de cor azulada, que originou o nome das cianobactérias, anteriormente denominadas "algas azuis") . Elas vivem no mar, na água doce e em meio terrestre úmido. Há espécies que possuem células isoladas e outras que formam colônias de diferentes formatos. A reprodução nas cianobactérias Nas cianobactérias unicelulares, a reprodução assexuada dá-se por divisão binária da célula. Nas espécies filamentosas, é comum a ocorrência de fragmentação do filamento, produzindo-se vários descendentes semelhantes geneticamente uns aos outros. A esses fragmentos contendo muitas células dá-se o nome de homogônios. |
As Arqueobactérias e seu Incrível Modo de Viver
Atualmente muitos autores consideraram oportuna a separação das Arqueobactérias (bactérias primitivas) das chamadas Eubactérias (bactérias verdadeiras).
Com base em estudos bioquímicos (sequências de RNA ribossômico, ausência de ácido murâmico na parede, composição lipídica da membrana), concluiu-se que há mais de 3000 M.a. teria ocorrido uma divergência na evolução dos organismos procariotas, tendo surgido duas linhagens distintas.
Até este momento não foi identificada recombinação genética neste grupo de organismos. O ramo que originou as Arqueobactérias teria, mais tarde, originado os eucariotas.
Considera-se que as arqueobactérias atuais pouca alteração sofreram, em relação aos seus ancestrais. Estes procariontes vivem em locais com condições extremamente adversas para outros seres vivos, provavelmente semelhantes às que existiriam na Terra primitiva.
As arqueobactérias podem ser divididas em três grandes grupos principais:
- Halófilas - vivem em concentrações salinas extremas, dezenas de vezes mais salgadas que a água do mar, em locais como salinas, lagos de sal ou soda, etc. A sua temperatura ótima de crescimento é entre 35 e 50ºC.
Estas bactérias são autotróficas, mas o seu mecanismo de produção de ATP é radicalmente diferente do habitual, pois utilizam um pigmento vermelho único - bacteriorrodopsina - que funciona como uma bomba de prótons (como os da fosforilação oxidativa nas mitocôndrias) que lhes permite obter energia;
- Metanogeneas - este grupo de bactérias foi o primeiro a ser reconhecido como único. Vivem em pântanos, no fundo dos oceanos, estações de tratamento de esgotos e no tubo digestivo de algumas espécies de insetos e vertebrados herbívoros, onde produzem metano (CH4) como resultado da degradação da celulose.
As reservas de gás natural que conhecemos são o resultado do metabolismo anaeróbio obrigatório e produtor de metano de bactérias deste tipo no passado. Algumas conseguem produzir metano a partir de CO2 e H2, obtendo energia desse processo.O gênero Methanosarcina consegue fixar azoto atmosférico, capacidade que se julgava única das eubactérias;
- Termoacidófilas -vivem em zonas de águas termais ácidas, com temperaturas ótimas entre 70 e 150ºC e valores de pH ótimo perto do 1. Na sua grande maioria metabolizam enxofre: podem ser autotróficas, obtendo energia da formação do ácido sulfídrico (H2S) a partir do enxofre, ou heterotróficas.
Reino Protista
A complexidade da célula eucariótica de um protozoário é tão grande, que ela - sozinha - executa todas as funções que tecidos, órgãos e sistemas realizam em um ser pluricelular complexo. Locomoção, respiração, excreção, controle hídrico, reprodução e relacionamento com o ambiente, tudo é executado por uma única célula, que conta com algumas estruturas capazes de realizar alguns desses papéis específicos, como em um organismo pluricelular.
Segundo a classificação dos seres vivos em cinco reinos (Whittaker – 1969), um deles, o dos Protistas, agrupa organismos eucariontes, unicelulares, autótrofos e heterótrofos. Neste reino se colocam as algas inferiores: euglenófitas, pirrófitas (dinoflagelados) e crisófitas (diatomáceas), que são protistas autótrofos (fotossintetizantes). Os protozoários são protistas heterótrofos.
A célula
A célula de um protista é semelhante às células de animais e plantas, mas há particularidades. Os plastos das algas são diferentes dos das plantas quanto à sua organização interna de membranas fotossintéticas. Ocorrem cílios e flagelos para a locomoção. A célula do protozoário tem uma membrana simples ou reforçada por capas externas protéicas ou, ainda, por carapaças minerais, como certas amebas (tecamebas). Os radiolários e heliozoários possuem um esqueleto intracelular composto de sílica. |  |
Os foraminíferos são dotados de carapaças externas feitas de carbonato de cálcio. As algas diatomáceas possuem carapaças silicosas.
Os protistas podem ainda ter adaptações de forma e estrutura de acordo com o seu modo de vida: parasita, ou de vida livre.
O citoplasma está diferenciado em duas zonas, uma externa, hialina, o ectoplasma, e outra interna, granular, o endoplasma. Nesta, existem vacúolos digestivos e inclusões.
Origem
 | Os protozoários constituem um grupo de eucariontes com cerca de 20 mil espécies. É um grupo diversificado, heterogêneo, que evoluiu a partir de algas unicelulares. Em alguns casos essa origem torna-se bem clara, como por exemplo no grupo de flagelados. Há registro fóssil de protozoários com carapaças (foraminíferos), que viveram há mais de 1,5 bilhão de anos, na Era Proterozóica. Grandes extensões do fundo dos mares apresentam espessas camadas de depósitos de carapaças de certas espécies de radiolários e foraminíferos. São as chamadas vasas. Ao lado: Microscopia eletrônica da carapaça presente externamente à célula de uma espécie de radiolário. |
Habitat
Os protozoários são, na grande maioria, aquáticos, vivendo nos mares, rios, tanques, aquários, poças, lodo e terra úmida. Há espécies mutualísticas e muitas são parasitas de invertebrados e vertebrados. Eles são organismos microscópicos, mas há espécies de 2 a 3 mm. Alguns formam colônias livres ou sésseis.
Fazem parte do plâncton (conjunto de seres que vivem em suspensão na água dos rios, lagos e oceanos, carregados passivamente pelas ondas e correntes). No plâncton distinguem-se dois grupos de organismos:
- fitoplâncton: organismos produtores (fotossintetizadores), representados principalmente por dinoflagelados e diatomáceas, constituem a base de sustentação da cadeia alimentar nos mares e lagos . São responsáveis por mais de 90% da fotossíntese no planeta.
- zooplâncton: organismos consumidores, isto é, heterótrofos, representados principalmente por protozoários, pequenos crustáceos e larvas de muitos invertebrados e de peixes.
Digestão
Nas espécies de vida livre há formação de vacúolos digestivos. As partículas alimentares são englobadas por pseudópodos ou penetram por uma abertura pré-existente na membrana, o citóstoma.

Já no interior da célula ocorre digestão, e os resíduos sólidos não digeridos são expelidos em qualquer ponto da periferia, por extrusão do vacúolo, ou num ponto determinado da membrana, o citopígio ou citoprocto.
Respiração
A troca de gases respiratórios se processa em toda a superfície celular.
Excreção
Os produtos solúveis de excreção podem ser eliminados em toda a superfície da célula. Nos protozoários de água doce há um vacúolo contrátil, que recolhe o excesso de água absorvido pela célula, expulsando-a de tempos em tempos por uma contração brusca. O vacúolo é, portanto, osmorregulador.
Classificação
A classificação dos protozoários baseia-se fundamentalmente nos tipos de reprodução e de organelas locomotoras.
A locomoção se faz por batimento ciliar, flagelar, por emissão de pseudópodos e até por simples deslizamento de todo o corpo celular. Em alguns ciliados há, no lugar do citoplasma, filamentos contráteis, os mionemas. Os pseudópodos, embora sendo expansões variáveis do citoplasma, podem se apresentar sob diferentes formas.
Na tendência moderna, os protozoários estão incluídos no Reino Protista, subdivididos em quatro filos:
Rizópodes ou Sacorníceos
São amebas (“nus”); radiolários e foraminíferos (têm carapaças com formas bastante vistosas, feitas de calcário ou de sílica - importantes indicadores da existência de jazidas de petróleo)
São marinhos, de água doce ou parasitas (Entamoeba histolytica). Têm um ou mais núcleos, vacúolos digestivos e vacúolos contráteis (apenas nos de água doce).
Os Rizópodes caracterizam-se por apresentarem pseudópodes como estrutura de locomoção e captura de alimentos. São projeções da célula, que se deforma toda, que encaminham a ameba para várias direções. O mecanismo que leva à formação dos pseudópodes está hoje razoavelmente esclarecido: na região de formação de uma dessas projeções, a parte viscosa do citoplasma se torna fluida, permitindo que o restante da célula flua nessa direção. Vários pseudópodos podem ser formados ao mesmo tempo, modificando constantemente a forma da ameba. Os pseudópodos, na ameba, não servem apenas para a locomoção. Também são utilizados para a captura de alimento: pequenas algas, bactérias, partículas soltas na água etc. Eles rodeiam o alimento e o englobam.
O vacúolo alimentar formado (também chamado de fagossomo) une-se a lisossomo e se transforma em vacúolo digestivo. Inicia-se a digestão, a partir de enzimas lisossômicas que atuam em meio ácido. Progressivamente, o conteúdo do vacúolo digestivo torna-se alcalino, até completar-se a digestão. As partículas digeridas atravessam a membrana do vacúolo, espalham-se pelo citoplasma e vão participar do metabolismo celular. Partículas residuais são expelidas da célula pela fusão da parede do vacúolo com a superfície da célula, em um processo inverso ao da fagocitose.
As amebas de vida livre que vivem em água doce apresentam vacúolo contrátil ou pulsátil para osmorregulação, eliminando o excesso de água que vai entrando no seu citoplasma (hipertônico), vindo do ambiente mais diluído (hipotônico).

Microscopia de um risópode.
Em condições desfavoráveis, por exemplo sujeita à desidratação, a Entamoeba produz formas de resistência, os cistos, com quatro núcleos no seu interior (partição múltipla).
A reprodução assexuada é por bipartição simples ou cissiparidade (mecanismo semelhante a mitose).
Dentre as amebas é importante a Entamoeba histolytica, que parasita o intestino humano, causando a disenteria amebiana ou amebíase.
Flagelados
Sua célula é alongada, podem ter um ou mais flagelos e em alguns há também pseudópodos. No gênero Trypanosoma há uma membrana ondulante que auxilia na locomoção. Próximo ao ponto de origem do flagelo, existe o cinetoplasto, organela que contém o DNA, capaz de se autoduplicar e que fica incluído no interior de uma longa mitocôndria de formato irregular que se estende ao longo da célula.
Existem flagelados de vida livre (Euglena – possuem clorofila e realizam fotossíntese; podem, também, nutrir-se de forma heterótrofa = zooflagelados), mutualísticos (Trichonympha, no intestino de cupins – fornecem a enzima celulase) e parasitas (Trypanosoma cruzi).
Nos coanoflagelados, há uma espécie de colarinho que serve para a captura de partículas alimentares; têm estrutura muito semelhante aos coanócitos, células típicas das esponjas.
Devido a isso, há teorias que sugerem uma relação filogenética entre coanoflagelados e esponjas.
 Coanoflagelado |  |
A reprodução é sexuada ou assexuada por divisão longitudinal.
Este filo tem muitos importantes parasitas humanos:
- Leishmania braziliensis: Causa a leishmaniose tegumentar ou úlcera de Bauru ('ferida brava'). Vive no interior das células da pele e é transmitida pelo mosquito-palha (birigui).
- Trypanosoma cruzi: Causa a doença de Chagas, comum em nosso país e na América do Sul é transmitida por percevejos popularmente conhecidos como barbeiros.
- Giardia lamblia: Causa a giardíase (intestinal).
- Trichomonas vaginalis: Causa a tricomoníase (no aparelho genital).
- Trypanosoma cruzi: Causa a doença de Chagas, comum em nosso país e na América do Sul é transmitida por percevejos popularmente conhecidos como barbeiros.
- Giardia lamblia: Causa a giardíase (intestinal).
- Trichomonas vaginalis: Causa a tricomoníase (no aparelho genital).
No intestino dos cupins e das baratas que comem madeira existem flagelados. Essa convivência é pacifica e caracteriza uma associação em que ambos os participantes são beneficiados (mutualismo). A madeira ingerida pelos insetos é digerida por enzimas produzidas pelos flagelados. Ambos aproveitam os produtos da digestão.
Esporozoários ou Apicomplexos: são todos parasitas
Não possuem orgânulos para locomoção.
São todos parasitas e apresentam um tipo de reprodução assexuada especial chamada de esporulação: uma célula divide seu núcleo numerosas vezes; depois, cada núcleo com um pouco de citoplasma é isolado por uma membrana, formando assim vários esporos a partir de uma célula
No ciclo vital apresentam alternância de reprodução assexuada e sexuada.
O principal gênero é o Plasmodium, com várias espécies causadoras da malária. O Toxoplasma gondii, causador da doença toxoplasmose, é de grande seriedade em mulheres grávidas até o terceiro mês.

Ciliados
É o grupo mais altamente especializado. Apresentam cílios, cirros e membranelas. Estas duas últimas estruturas resultam da concrescência (união) de muitos cílios. Entre eles estão os protozoários “gigantes” como os paramécios (Paramecium) muito usados em estudos; aqui estão os protozoários de organização mais complexa. Os paramécios deslocam-se muito mais rapidamente que os flagelados e as amebas por causa dos inúmeros cílios que se projetam da parede do corpo. A maioria é de vida livre.
Além de orgânulos especializados, possuem dois núcleos: macronúcleo (funções vegetativas) e micronúcleo (funções genéticas: hereditariedade e reprodução); apresentam extremidades anterior e posterior; na membrana, a entrada do alimento se dá pelo citóstoma e a saída de resíduos pelo citopígio (= citoprocto).
Possuem dois vacúolos pulsáteis que funcionam alternadamente efetuando a regulação osmótica e possivelmente a expulsão de toxinas. Cada vacúolo possui canais que recolhem a água celular, encaminhando-a para um reservatório que efetua a sua expulsão da célula.
Trocas gasosas e excreção, como nos demais protozoários, ocorre pela superfície da célula. A reprodução assexuada, como na ameba e na euglena, ocorre por divisão binária.
A reprodução sexuada por conjugação consiste no pareamento de dois paramécios, com fusão das membranas e em seguida troca de material genético dos micronúcleos. Depois os paramécios se separam e se reproduzem assexuadamente por cissiparidade.
